|
reklama
| Systémová enzymoterapia a jej vplyv na funkciu imunitného systému |
|
prof. MUDr. Milan Buc DrSc. |
| SANQUIS č.58/2008, str. 100 |
 |
zpět na výběr odborných článků |
Enzýmy sú najvýznamnejšími biokatalyzátormi, v živých organizmoch umožňujú a regulujú všetky biochemické procesy. Rozdeľujeme ich do 6 skupín, z ktorých medicínsky najvýznamnejšia je skupina hydroláz. Hydrolázy štiepia zložité biologické látky, vrátane bielkovín. Táto ich vlastnosť pritiahla záujem lekárov o ich využitie v liečbe rôznych chorôb.
|
|
Skutočne, záznamy o enzymoterapii siahajú do dávnoveku – využívali sa pri liečbe rán a zápalov aj mnohých celkových chorôb. Aj modernej medicíne nie je enzymoterapia neznáma. Enzýmy, resp. ich zmesi sa využívajú pri poruchách trávenia, k trombolýze akútnych infarktov a ďalších trombotických a embolických príhod; využíva sa aj lokálna aplikácia.
Systémová enzymoterapia (SET) predstavuje liečebný postup, pri ktorom sa chorým podávajú, perorálne, vysoké dávky cielene zostavených kombinácií proteolytických enzýmov živočíšneho pôvodu (pankreatický extrakt, trypsín, chymotrypsín) a rastlinného pôvodu (bromelaín, papaín). Rozmach systémovej enzymoterapie umožnil najmä dôkaz, že črevnou stenou môže prechádzať značná časť perorálne podaných enzýmov nielen vo forme fragmentov, ale aj v pôvodnej makromolekulovej a funkčne aktívnej forme. (4)
Proteinázy, po požití vo forme gastrorezistentných tabletiek, sa z oblasti tenkého čreva vstrebávajú do krvného obehu. Tu sa nadväzujú na antiproteinázy ( α2-makroglobulín, α1-proteinázový inhibítor), čím sa znižuje ich antigénnosť. V komplexoch „proteináza – antiproteináza“ sú navyše chránené pred rozpoznaním bunkami imunitného systému a v nepoškodenej forme sa dostávajú do oblastí postihnutých zápalom. V tejto forme sa tiež realizuje väčšina ich rozhodujúcich farmakologických účinkov, ako je tlmenie zápalu, odstraňovanie imunokomplexov a modulácia imunitnej odpovede. (13)
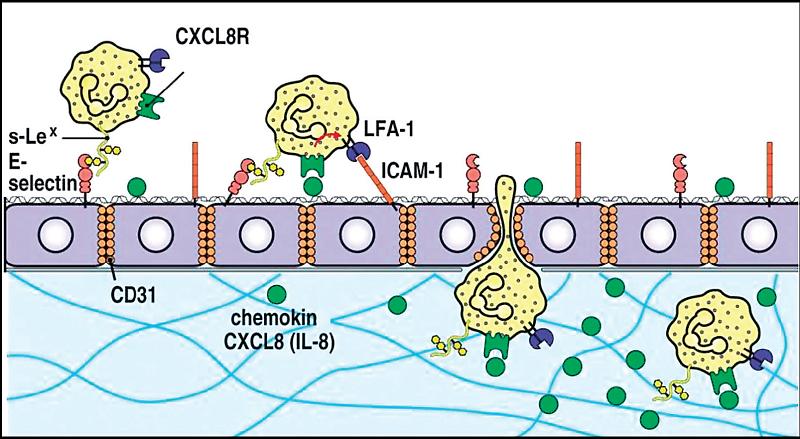
Protizápalové pôsobenie trypsínu, chymotrypsínu, papaínu a bromelaínu, ako aj ich kombinácie, sa dokázalo v početných experimentálnych prácach. Enzýmy odbúravajú poškodené bunky a nekrotický materiál a inaktiváciou mediátorov a toxických produktov zmenšujú edém a zmierňujú bolesť. Znížením expresie adhezívnych molekúl sa znižuje prechod leukocytov stenou kapilár, čím sa obmedzuje ich prísun do zápalového ložiska (7) (obr. 1).
|
|
|
| Schéma 1: Interakcia leukocytov pri jeho prechode do extravaskulárneho prostredia (Janeway et al. 2005, upravené) Leukocyty majú v svojich membránach adhezívne molekuly, pomocou ktorých interagujú s adhezívnymi molekulami v membránach endotelových buniek. Prvé interakcie, po tom ako leukocyt získal chemotaktický podnet, napr. v podobe IL-8, sa odohrávajú na úrovni selektínov, kde E-selektín (CD62E) sa viaže na S-Lex (sialyl Lewis), čo spôsobí, že leukocyt začne po povrchu endotelu „skákať“. Súčasne sa v jeho membráne exprimuje iná skupina adhezívnych molekúl, integríny. Z nich najvýznamnejší je LFA1-1, ktorý sa viaže s ICAM-1 na endotele. Táto interakcia už spôsobí, že leukocyt zostane na povrchu endotelu pevne prichytený, mení svoj tvar z okrúhleho na plochý a následne štrbinami medzi endotelovými bunkami sa preplazí (diapedéza) do extravaskulárneho prostredia.
|
|
Na druhej strane enzýmy zase zvyšujú funkčnú aktivitu neutrofilov, čo sa prejavuje zvýšením hodnôt respiračného vzplanutia a zvyšuje sa aj mikrobicídna aktivita makrofágov. (1) Podobne enzýmy môžu aktivovať T-lymfocyty a tak indukovať mechanizmy špecifickej imunity. (16, 18)
Celkové príznaky zápalu ovplyvňuje SET zásahom do porušenej cytokínovej siete. Cytokíny sa v plazme viažu (podobne ako proteinázy) na antiproteinázy (najmä na 2- makroglobulín). Väzba cytokínu na samotnú antiproteinázu nie je pevná a cytokín po svojom uvoľnení môže získať svoju aktivitu znovu. Pokiaľ však dôjde k väzbe cytokínu na 2-makroglobulín, na ktorý sa už predtým nadviazala proteináza, vzniká pevná väzba, ktorá cytokín inaktivuje a celý takto vzniknutý komplex (proteináza – antiproteináza – cytokín) sa rýchlo eliminuje makrofágmi pečene (Kupfferove bunky) a sleziny. (10) Proteinázy SET takto urýchujú elimináciu patologicky zvýšených hladín niektorých cytokínov.
Znížením hladiny prozápalových cytokínov (TNF, IL-1 a IL-6) proteinázy zabraňujú nadmernej a hlavne dlhodobej aktivácii buniek a bránia tak prechodu zápalu do chronicity. (15) Podobne dochádza aj k zníženiu hladín TGF-β , čím sa bráni progredujúcej fibróze, ktorá sa vďaka nemu pri chronickom zápale rozvíja. (5)
Urýchlenie odbúrania prozápalových cytokínov znižuje aj pravdepodobnosť protrombogénnej konverzie endotelu, čo sa spolu so znížením koncentrácie fibrinogénu a bradykinínu, znížením agregácie trombocytov a zvýšením koncentrácie plazminogénu spolupodieľa na fibrinolytickom, resp. antitrombogénnom účinku proteináz. Celkovo možno konštatovať, že enzýmy zápal nepotláčajú, ale optimalizujú jeho priebeh do vhodnej intenzity a rozsahu a bránia prechodu do chronickej formy. (14)
|
|
Redukcia edému spolu s antitrombogénnym pôsobením SET zlepšuje aj reologické vlastnosti krvi. Podpora mikrocirkulácie výrazne ovplyvňuje regeneráciu tkaniva, pretože obnoviteľnosť fyziologickej funkcie ložiska, ktoré je postihnuté zápalom, priamo závisí aj od doby, počas ktorej je okolitá mikrocirkulácia blokovaná. Zlepšenie reologických vlastností krvi a lymfy navyše priaznivo ovplyvní prestup antibiotík a chemoterapeutík do tkaniva. (13, 14)
Komplexy antigénu s protilátkou (imunokomplexy) vznikajú v organizme pri každom stretnutí protilátky so solubilným antigénom. Imunokomplexy sa vo väčšine prípadov vychytávajú a odstraňujú bunkami mononukleárno-fagocytového systému.
Za určitých okolností tvorba imunokomplexov môže mať patologické dôsledky, a to vtedy, ak sa vytvoria podmienky na ich deponovanie do tkaniva; ukladajú sa najmä do stien kapilár, glomerulov a synovie kľbov a vyvolajú tu zápal. Klinicky sa prejaví ako vaskulitída, glomerulonefritída alebo artritída, príp. iný zápal. (3)
V terapii imunokomplexového zápalu sa ako podporná liečba odporúča aj systémová enzymoterapia. Mechanizmus účinku sa prejavuje na viacerých úrovniach. Enzýmy už v nízkych koncentráciách znižujú hladinu imunokomplexov. (13, 14) Pôsobením enzýmov na imunokomplexy nadviazaný C3b- -fragment komplementového systému sa transformuje na inaktivovaný iC3b. Pre tento fragment majú makrofágy a neutrofily receptory (CR1, CR3), čím sa uľahčí pohltenie komplexov „iC3b – imunokomplex“ a následne aj ich degradácia.
Postupný pokles koncentrácie imunokomplexov v krvi a tkanivách umožňuje aj obnovu clearanceovej schopnosti mononukleárno- fagocytového systému, a to tým skôr, že enzýmy zvyšujú aktivitu fagocytov. (1, 16, 18)
Veľmi citlivá na pôsobenie enzýmov je aj druhá konštantná doména (CH2) IgG; jej alteráciou sa zabráni väzbe prvej zložky klasickej dráhy aktivácie komplementu, C1q, na protilátku a tým následne jeho aktivácii. (9) Výsledkom aktivity enzýmov je, že sa zníži tvorba vazoaktívnych a chemoatrahujúcich fragmentov (C5a > C3a > C4a) a zápal sa utlmí. (2, 11) Aktivácia T-lymfocytov je kľúčovým momentom pri indukcii imunitnej odpovede antigénmi proteínovej povahy. Je známe, že nato, aby antigén navodil imunitnú odpoveď, sa musí najprv opracovať v bunkách prezentujúcich antigén (APC = dendritové bunky, makrofágy). Tieto antigén pohltia, degradujú na menšie peptidy a nadviažu na vlastné HLA-molekuly druhej triedy (HLA-DR, -DQ, -DP). Po väzbe peptidu sa HLA-molekuly presúvajú do membrány bunky a takto „vystrčia“ nadviazaný peptid do extracelulárneho prostredia; hovoríme o prezentácii antigénu. Takto prezentovaný peptid v ďalšom procese imunitnej odpovede rozpoznávajú pomocné T-lymfocyty (TH).
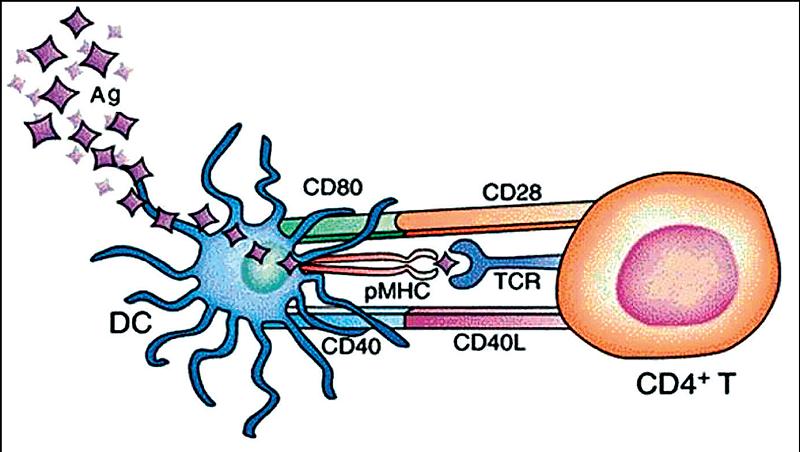
Proces rozpoznávania prebieha v dvoch fázach. V prvej fáze T-lymfocyty svojím špecifickým receptorom (TCRαβ ) rozpoznávajú prezentovaný peptid; súčasne dochádza k interakcii medzi HLA-molekulami na bunkách prezentujúcich antigén a diferenciačnými CD4-antigénmi TH-lymfocytov. Touto interakciou sa zabezpečí prvý signál potrebný pre ich aktiváciu. Druhý signál T-lymfocyty získavajú z interakcie kostimulačných molekúl – na strane T-lymfocytov je to molekula CD28 a na strane APC diferenciačný antigén CD80, resp. CD86 (obr. 2). Až integrácia signálov, aj z TCR aj z kostimulačných dráh, vedie k úplnej aktivácii T-lymfocytu, jeho proliferácii a napokon k vytvoreniu klonu buniek, ktoré dokážu špecificky reagovať s antigénom, ktorý imunitnú odpoveď vyvolal. (2, 11)
|
|
|
Schéma 2: Rozpoznávanie antigénu pomocným T-lymfocytom Antigén (Ag) po vniknutí do organizmu zachytia a spracujú dendritové bunky (DC). Spracovanie pozostáva z rozštiepenia proteínu na menšie fragmenty, peptidy, ktoré sa nadviažu do žliabku molekúl hlavného histokompatibilného komplexu (pMHC). Tieto peptidy rozpoznajú pomocné T-lymfocyty (CD4+ T) svojim antigénovým receptorom (TCR). Súčasne prebehnú kostimulačné interakcie medzi molekulami CD80 a CD28, resp. CD40 a CD40L.
Enzýmy používané v SET špecificky znižujú množstvo molekúl CD4 a CD80 v membránach kooperujúcich buniek, čím značne ovplyvňujú prahovú aktiváciu T-lymfocytov. (6, 7) Na to, aby sa T-lymfocyt aktivoval, musí na APC rozpoznať približne 300 rovnakých peptidov viazaných na svoje HLA-molekuly. Počet rozpoznávaných peptidov potrebných na aktiváciu T-lymfocytu závisí aj od afinity jeho antigénového receptora a množstva adhezívnych molekúl. Vysoký počet adhezívnych molekúl umožní T-lymfocytu, aby sa aktivoval aj pri rozpoznávaní menšieho počtu prezentovaných peptidov, a naopak, nižší počet adhezívnych molekúl si vyžaduje, aby T-lymfocyt rozpoznal väčší počet prezentovaných peptidov.
Práve tento princíp sa uplatňuje pri fyziologickom posilňovaní imunitnej odpovede. Viaceré cytokíny, najmä interferón- gama (IFN-γ ) a interleukín 2 (IL-2), ktoré vznikajú v procesoch imunitnej odpovede, zvyšujú expresiu HLA-molekúl druhej triedy. Podobne, aktivovaný T-lymfocyt zvyšuje v svojej membráne množstvo CD4-molekúl. (2, 11)
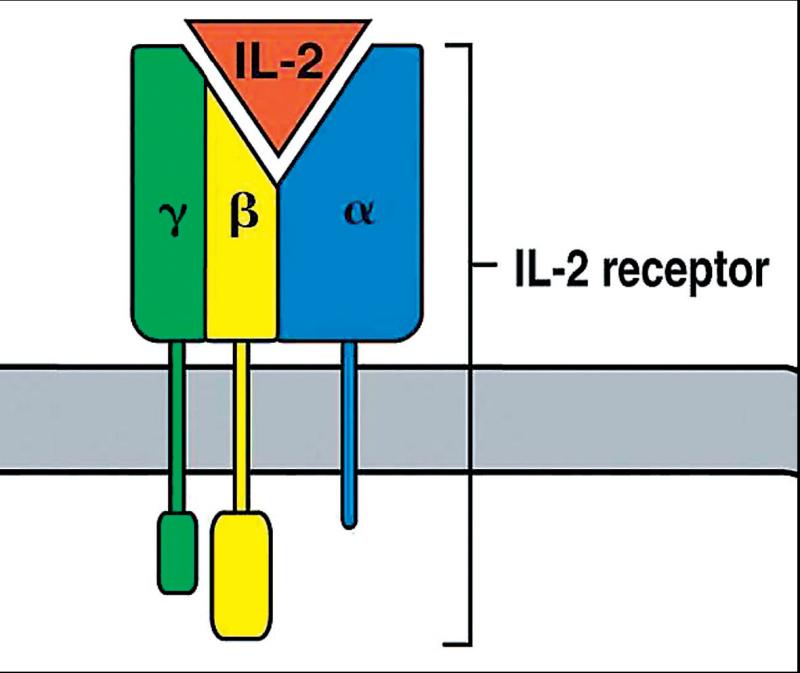
Enzýmy pri SET pôsobia opačne – znižovaním počtu adhezívnych a kostimulačných molekúl zvyšujú prah aktivácie T-lymfocytov. (6, 12) Významné v tejto súvislosti môže byť najmä znižovanie expresie receptora pre IL-2. T-lymfocyt po svojej aktivácii začne syntetizovať IL-2 a exprimovať receptor receptor pre tento cytokín. IL- 2 autokrinným spôsobom podnieti proliferáciu T-lymfocytov a ich klonovú expanziu. Receptor pre IL-2 (IL-2R) sa skladá z troch reťazcov – α (p55 = CD25), β (p75 = CD122) a γ (p64). Rôzna kombinácia týchto receptorov umožňuje vznik odlišných receptorov. Kombinácia reťazcov β a γ je potrebná pre prenos signálu, avšak pre vznik vysokoafinitného receptora je nevyhnutná aj prítomnosť α-reťazca. IL-2R skladajúci sa iba z reťazcov α a β vytvára nízkoafinitný receptor (Kd = 10-9 M), zatiaľ čo receptor, ktorý sa skladá zo všetkých troch reťazcov, je vysokoafinitný (Kd = 10-11 M) (obr. 3). B-lymfocyty majú približne 1000 vysokoafinitných receptorov, T-lymfocyty 10krát viac. (2, 11) Zníženie expresie vysokoafinitného receptora enzýmami imunitnú odpoveď takto utlmí. Druhý mechanizmus, ktorým enzýmy pri SET utlmujú imunitnú odpoveď, je znižovanie syntézy cytokínov, ktoré sú dôležité v iniciácii či udržiavaní imunopatologických procesov. (15) Uvedené vysvetlenie pôsobenia enzýmov na zložky a funkciu imunitného systému naznačuje možnosť ich využitia v medicínskej praxi. Indikačné pole SET je najmä v liečbe zápalových procesov a autoimunitných chorôb.
V liečbe zápalových procesov sa priaznivý účinok SET udáva pri terapii zápalov horných a dolných dýchacích ciest (akútna sinusitída, hnisavé bronchitídy, bronchiolitídy a peribronchitídy), adnexitíd, prostatitíd, cystitíd a cystopyelitíd, tromboflebitíd. Z autoimunitných chorôb sa SET používa pri liečbe reumatických chorôb a pri roztrúsenej skleróze (sclerosis multiplex); z imunokomplexových chorôb je liečba SET indikovaná najmä pri vaskulitídach a glomerulonefritídach. Napokon sa SET používa aj pri liečbe lymfedému.
|
|
|
Schéma vysokoafinitného receptora pre interleukín 2 Vysokoafinitný receptor pre interleukín 2 (IL-2) sa skladá z reťazcov α,β a γ; nízkoafinitnému receptoru naproti tomu chýba α-reťazec, tvoria ho iba reťazce β a γ. 1 Respiračné vzplanutie je jav, kedy pri metabolickej aktivácii fagocytu dochádza k zvýšenej, viac ako 100násobnej, spotrebe kyslíka.
Literatúra:
Biziulevicius GA, Where do the immunostimulatory effects of oral proteolytic enzymes („systemic enzyme therapy“) come from? Microbial proteolysis as a possible starting point. Med Hypotheses 2006; 67: 1386-8.
Buc, M.: Imunológia. Bratislava, Veda 2001, 463 s.
Buc, M.: Autoimunita a autoimunitné choroby. Bratislava, Veda 2005, 491 s.
Castell JV, Friedrich G, Kuhn CS, Poppe GE, Intestinal absorption of undegraded proteins in men: presence of bromelain in plasma after oral intake. Am J Physiol. 1997; 273: 139-46.
Desser L, Holomaňová D, Závadová E, Pavelka K, Mohr T, Herbáček I, Oral therapy with proteolytic enzymes decreases excessive TGF-beta levels in human blood. Cancer Chemother Pharmacol 2001; 47: S10-S15.
Engwerda ChR, Andrew D, Ladhams A, Tracey L, Mynott ML, Bromelain modulates T cell and B cell immune responses in vitro and in vivo. Cellular Immunol 2001; 210: 66-75.
Hale LP, Greer PK, Sempowski GD, Bromelain treatment alters leukocyte expression of cell surface molecules involved in cellular adhesion and activation. Clin Immunol 2002; 104: 183-90. Hale LP, Greer PK, Trinh CT, James CL, Proteinase activity and stability of natural bromelain preparations. Int Immunopharmacol 2005; 5: 783-93.
Heinz HP, Loos M, Activation of the first component of complement, C1: comparison of the effect of 16 different enzymes on serum C1. Immunobiol 1982; 165: 175-85.
James K, Interactions between cytokines and alfa2-macroglobulin. Immunol Today 1990; 11: 163- 6.
Janeway Ch, Travers P, Walport M, Shlomchik M, Immunobiology. The immune system in health and disease. 6th ed. New York, Tokyo: Churchill Livingstone /Garland Publishing, 2005: 732.
Manhart N, Akomeah R, Bergmeister H et al. Administrations of proteolytic enzymes bromelain and trypsin diminish the number of CD4+ cells and the interferon-γ response in Peyer´s patches and spleen in endotoxemic Balb/c mice. Cell Immunol 2002; 215: 113-9.
Nouza K, Olejár T, Nouza M, Proteinázy v regulaci a modulaci imunity. Klin imunol Alergol 1999; 3: 22-7.
Nouza K, Nouza M, Zánět optimalizující a imunonormalizační účinky kombinací proteolytických enzymù. Alergie 2002; 4: 326-9.
Onken JE, Greer PK, Calingaert B, Hale LP, Bromelain treatment decreases secretion of proinflammatory cytokines and chemokines by colon biopsies in vitro. Clin Immunol. 2008; 126: 345-52.
Ossovskaya vs, Bunnet nw, Protease-Activated Receptors: Contribution to Physiology and Disease Physiol. Rev. 2004; 84: 579-621.
Rose B, Herder C, Löffler H, Meierhoff G, Schloot NC, Walz M, Martin S, Dose-dependent induction of IL-6 by plant-derived proteases in vitro. Clin exp Immunol 2006; 143: 85-92.
Trejo J., Protease-activated receptors: new concepts in regulation of G protein-coupled receptor signaling and trafficking. J Pharmacol exp Ther 2003; 307: 437-42.
Celý článek ve formátu pdf naleznete zde.
|
|
|
|
obsah čísla 58 |
 |
ročník 2008 |
|
témata |
|
|